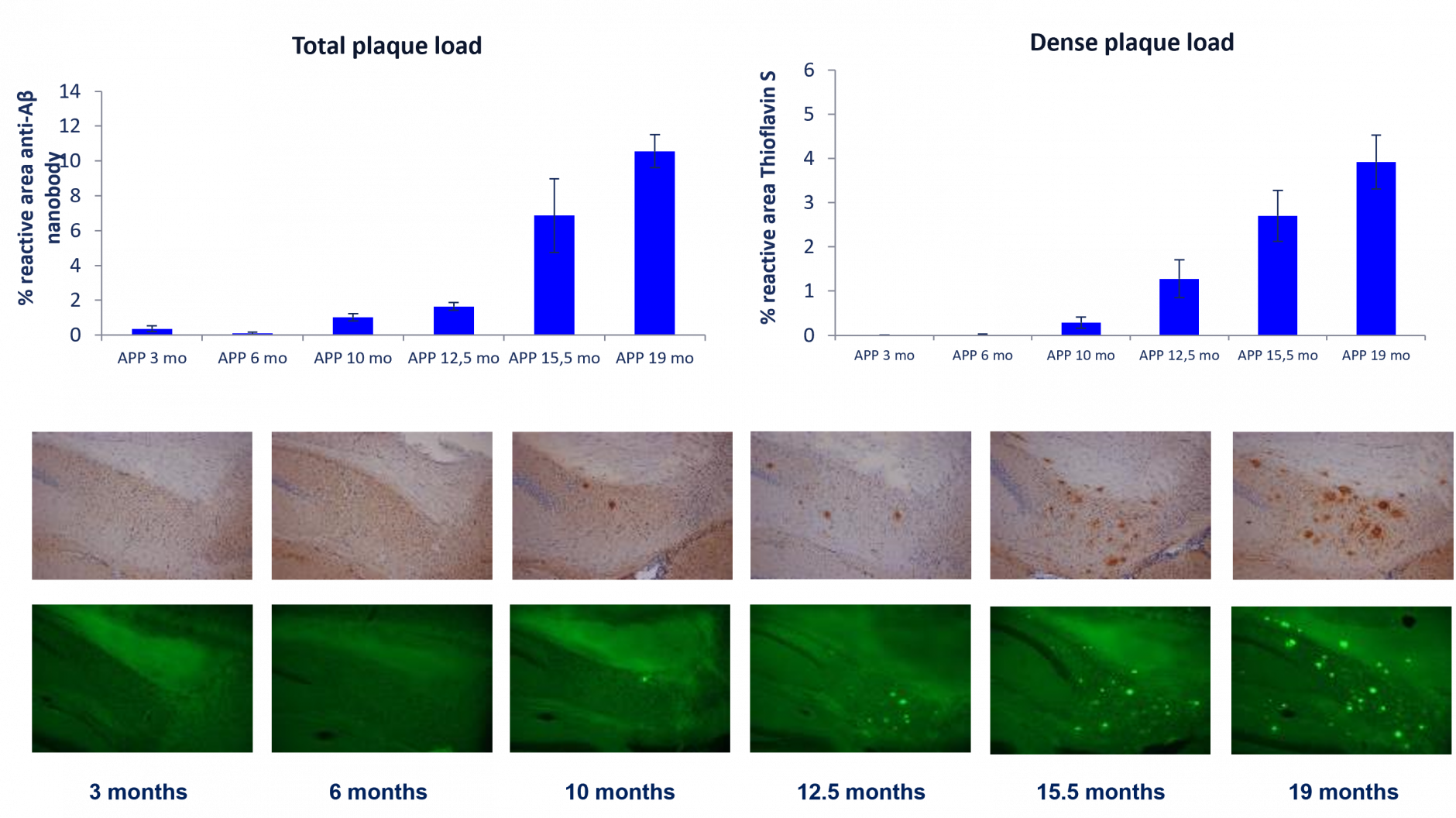
Progressive total plaque load (an anti-Aβ antibody) and dense plaque load (Thioflavin S) in the subiculum as measured by IHC (mean ± SEM, N = 5-9).
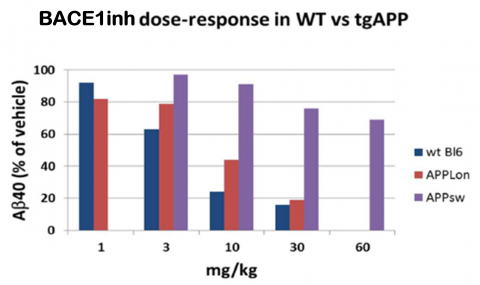
The London mutation [V717] is a preferred model for anti-BACE1 approaches. The Swedish mutation might result in an aberrant subcellular localization of APP (Jacobsen et al., 2014).
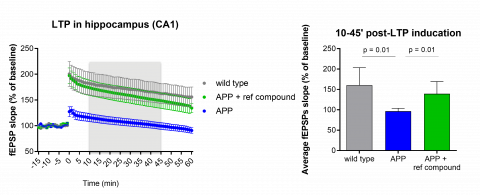
Long term potentiation deficit (induced by a high frequency stimulation) in the CA1 region of the hippocampus in 8 month old APP[V717] animals and age-matched littermates can be restored with a therapeutic compound (mean ± SEM, n = 5 per group).
We have a more extensive datapackage available. Contact us at cro[at]remynd.com to set up a meeting and discuss how we translate your mechanism of action into an effective study design.
Describing papers:
Therapeutic intervention papers:
reMYND, Bio-Incubator (Wetenschapspark)
Gaston Geenslaan 1
BE-3001 Leuven (Heverlee)
Belgium
RPR Leuven, 0476.910.101
BTW/VAT: BE0476.910.101
T +32 16 75 14 20
F +32 16 75 14 21
info@remynd.com
