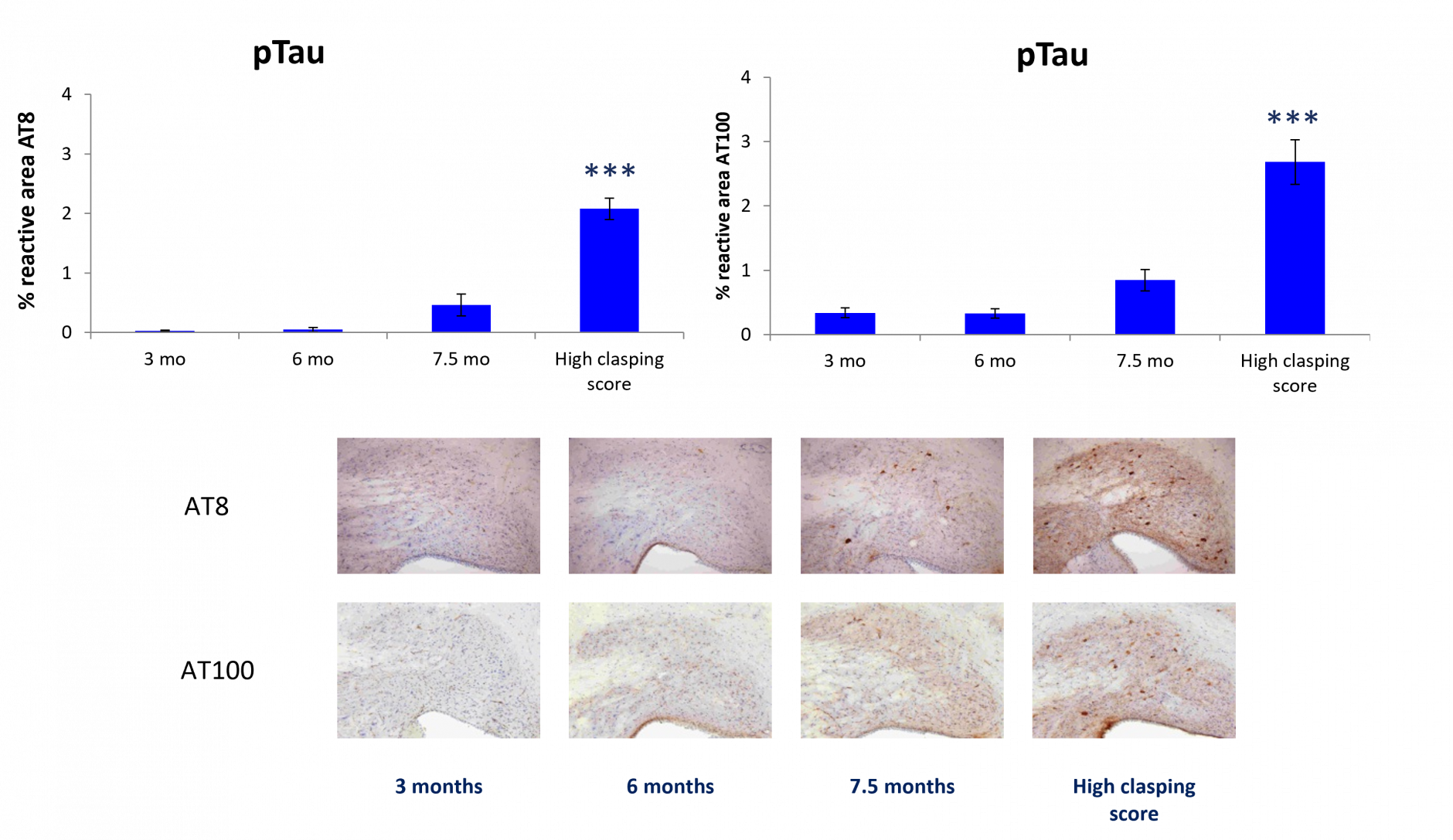
Age dependent increase of Tau pathology (AT8 = pSer202 pThr205 Tau, AT100 = pThr212/pSer214 Tau) in the interposed cerebellar nulceis, anterior and posterior part (LAT/Int/AP) as measured by IHC (mean ± SEM, n = 15). Statistics: One-way ANOVA
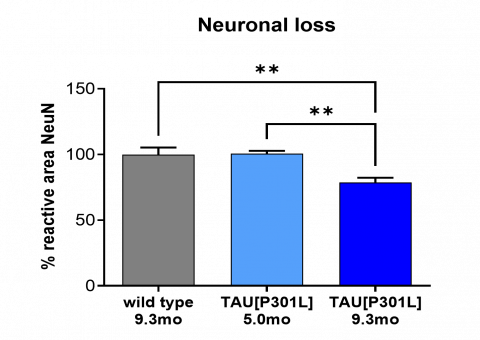
Progressive neuronal loss (NeuN) in the subthalamic nucleus and zona inserta (STH/ZI, n = 14, 15 and 28 respectively), as measured by IHC. Statistics: Two-way ANOVA, Tukey’s multiple comparison
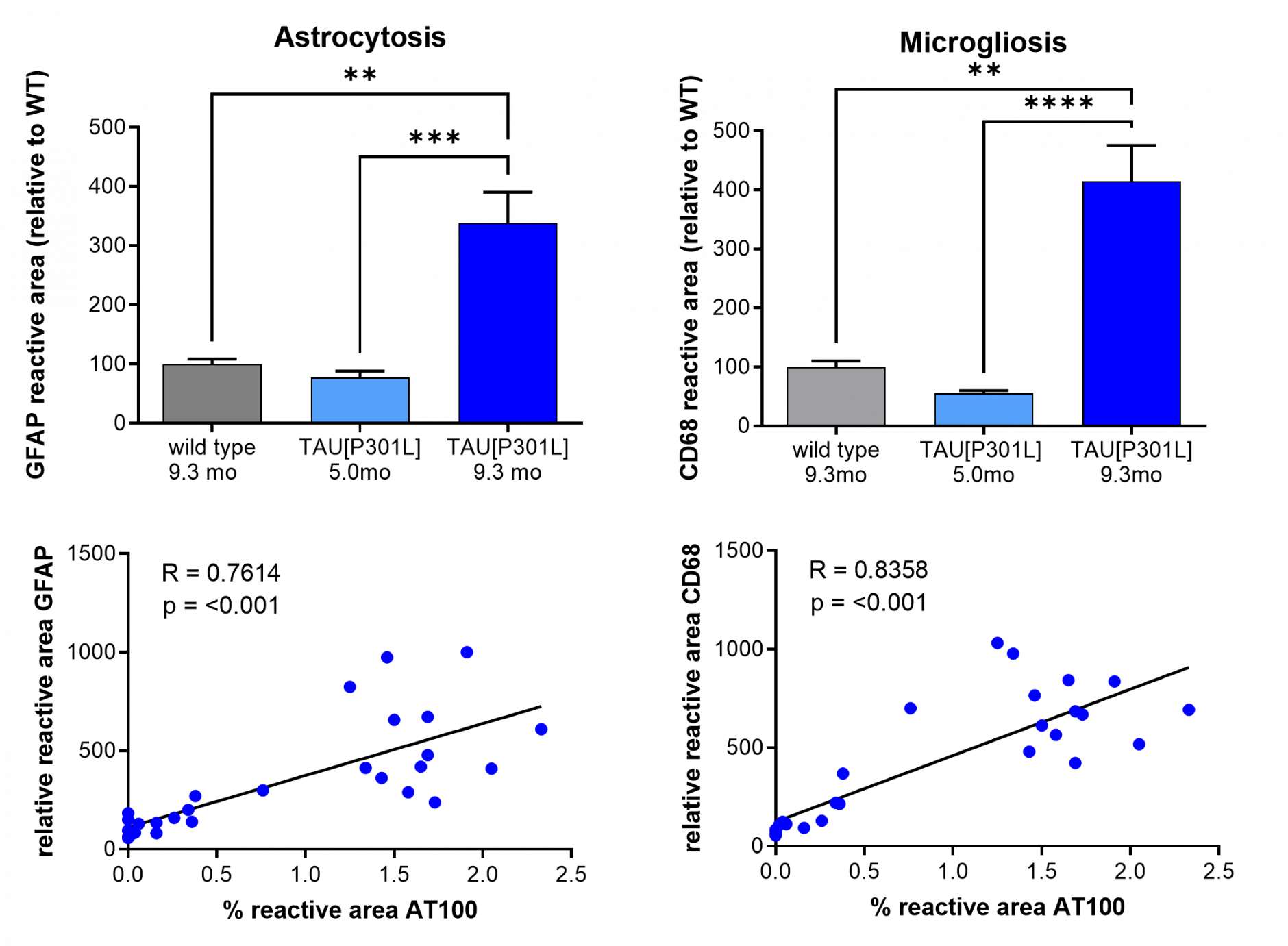
Progressive astrocytosis (GFAP) and microgliosis correlates with Tau pathology in the interposed cerebellar nulceis, anterior and posterior part (LAT/Int/AP) as measured by IHC (mean ± SEM, n = 14, 15 and 28 respectively). The correlation graphs are only for 9.3 months of age. Statistics: Two-Way ANOVA, Tukey’s multiple comparison
We have a more extensive datapackage available. Contact us at cro[at]remynd.com to set up a meeting and discuss how we translate your mechanism of action into an effective study design.
Describing papers:
Therapeutic intervention papers:
reMYND, Bio-Incubator (Wetenschapspark)
Gaston Geenslaan 1
BE-3001 Leuven (Heverlee)
Belgium
RPR Leuven, 0476.910.101
BTW/VAT: BE0476.910.101
T +32 16 75 14 20
F +32 16 75 14 21
info@remynd.com
